
Expresión de los genes. Síntesis de proteínas
• Teoría de un gen- un enzima.
• Del DNA a la proteína. Expresión del mensaje genético
NOTA. El fundamento de esta lección es comprender cómo desde el gen se llega al carácter, es decir,
• Transcripción. Síntesis del RNA
o Maduración del m-RNA transcrito en las células eucariotas
o El código genético: carácterísticas y desciframiento
o Traducción o Biosíntesis de las proteínas
(Adaptación de los Aa a sus correspondientes
t-RNA) (Iniciación de la síntesis de la cadena
polipeptídica)(Alargamiento) (Terminación)
o Regulación de la expresión de los genes en procariotas ( entender al menos la regulación del operón lac, daremos material complementario)
o Aproximación general a la regulación de la misma en eucariotas
1.-Un gen-una enzima
 |
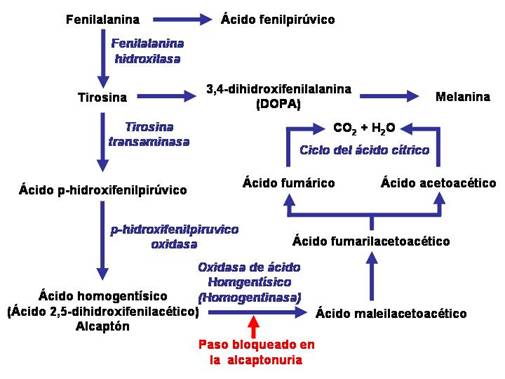
1901. Garrod, había observado ciertas enfermedades humanas hereditarias, que por tanto no tenían procedencia microbiológica. Una de ellas, la alcaptonuria, se caracterizaba por artritismo y ennegrecimiento de los cartílagos y orina. Estudiando la genealogía de los individuos, se pudo deducir que la alcaptonuria sólo aparecía cuando las dos informaciones progenitoras lo indicaban. Se trataba de una información biológica anormal, recesiva respecto a la correcta. |
En 1930, Beadle estudiando el color de ojos de Drosophila, formuló la hipótesis de que cada uno de los colores de los ojos de los mutantes, procedían de un cambio en una enzima.
El término información biológica hereditaria para un carácter fue sustituido por el de "gen". Un gen puede definirse como un fragmento de ADN que tiene la información para un determinado carácter. Ocupa en el filamento del ácido nucléico una posición fija denominada locus. En un mismo locus puede haber más de un tipo de gen y cada uno de los diferentes genes que existen en el locus se denomina alelo. En el caso anterior se trat de un locus con dos alelos, el normal y al alcaptonuria. Mediante análisis se averiguó que el motivo del ennegrecimiento de la orina y cartílagos se debía a la presencia del ácido homogentísico. Se supuso que en los individuos sanos esta sustancia era transformada en otras y desaparecía. Se pasó así del paralelismo un gen: un carácter a un gen-sustancia.
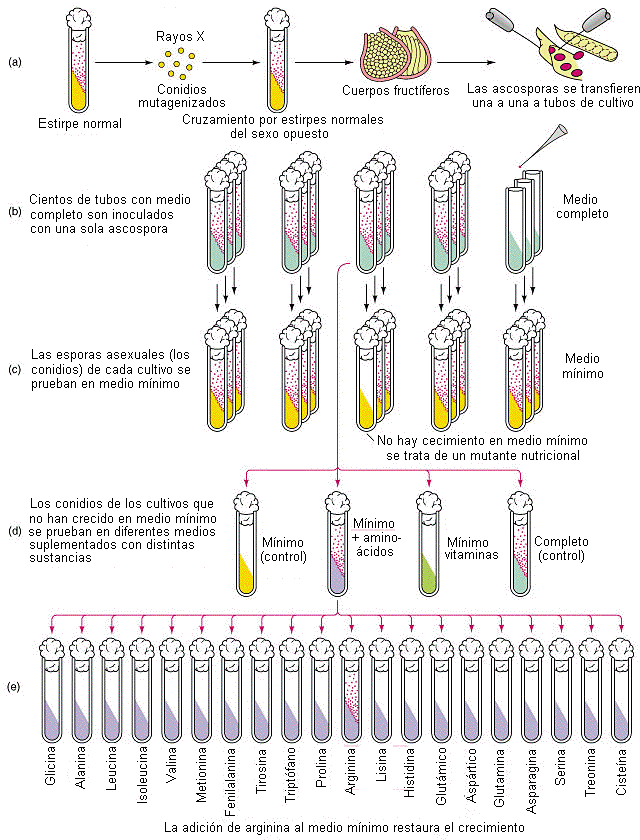
En 1948, Beadle y Tatum hicieron experimentos con Neurospora crassa, que necesita para vivir sustancias minerales, fuente de C y biotina. Mediante radiación con uv aparecieron mutantes que sólo sobrevivían si se les añadía arginina. Habían perdido, la capacidad de sintetizarla. Otros necesitaban arginina o citrulina, otros arginina o cirtrulina u ornitina. Se dedujo que la síntesis de arginina debía segur la siguiente vía metabólica:
| Sustrato | ornitina | citrulina | arginina | proteínas |
Extraído de http://www.ucm.es/info/genetica/grupod/Genenzima/Genenzima.htm
Mediante análisis bioquímicos se pudo confirmar que los diferentes mutantes carecían de determinadas enzimas y que en cada uno de ellos aumentaba mucho uno de los componentes de la cía de la arginina. Al faltar un enzima, el metabolismo queda bloqueado en aquella sustancia sobre la cual debía actuar. Se estableció así un paralelismo entre genes y enzimas, denominándose esta hipótesis: "teoría de un gen- una enzima" Al alterarse la secuencia de nucleótidos de un gen falta una enzima. Las enzimas controlan las sustancias y por ellas las características de los organismos.
Un hongo normal tiene todas las enzimas normales y un mutante no tiene suficiente con el sustrato normal, pero si se añade al medio el nutriente que necesita, sobrevivirá ya que tienen el resto de las enzimas.
2.-Del DNA a la proteína
Podemos decir, de forma general, que existen tantos enzimas como reacciones metabólicas se producen en la célula y que su ausencia provoca que una reacción metabólica no pueda efectuarse.
Crick propuso la "hipótesis de la colinearidad" por la que se establece una correspondencia entre la secuencia de nucleótidos y la de aminoácidos de la enzima que el gen codifica.
Mediante diferentes experimentos se llegó a la conclusión de que en el mecanismo por el cual se pasaba de una secuencia a la otra se podían diferenciar dos procesos : uno que se realizaba en el núcleo y otro en los ribosomas. En el núcleo se pasa de una secuencia de bases nitrogenadas de un gen a una secuencia e bases nitrogenadas complementarias pertenecientes a un ARNm; es la Transcripción. En los ribosomas se pasa de una secuencia de ribonucléotidos del ARNm a una secuencia de aminoácidos por la Traducción.


Síntesis proteica
Antes que nada, vamos a recordar que hay tres tipos de ARN, que van a facilitar la síntesis.
El ARNm
Diferencias entre la transcripción y la replicación
1.-La transcripción es selectiva: sólo se transcriben algunas regiones del DNA (los genes)
2.-Cuando se transcribe un gen, se copia una sola hebra del ADN, la hebra molde, con lo que la secuencia del RNA sintetizado es equivalente (con la sustitución de T por U a la de la hebra que no se transcribe, llamada hebra codificadora. Esto no quiere decir que todas las secuencias codificadoras se encuentren en la misma cadena del DNA: hay genes que se transcriben de una hebra y otros en otras regiones del DNA, que lo hacen de la contraria.
3.-La transcripción es reiterativa: un gen puede transcribirse muchas veces, mientras que la replicación sólo ocurre una vez antes de la división celular.
1.-Transcripción : ADN----ARN
Hay que resaltar que existen tres tipos de RNA poli en eucariotas ( en procariotas sólo 1).. La I que cataliza la síntesis de ARNr. La II que cataliza la síntesis del precursor del ARNm y la III que cataliza la de RNAt, RNAr y RNAu.
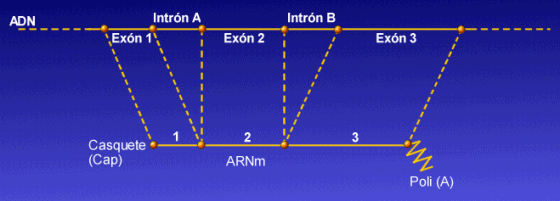
En eucariotas, los genes están fragmentados, de forma que siempre es necesario un proceso de maduración en el que se eliminen las secuencias sin sentido o intrones y se empalmen las secuencias con sentido o exones. Excepcionalmente hay genes, como los de las histonas que no presentan intrones,
Intrón: secuencia de bases que se transcriben pero no se traducen
Exón: secuencia de bases que se transcriben y se traducen.

Se piensa que los genes del primitivo antecesor debían de tener intrones. Las bacterias los habrán perdido por selección natural, porque se dividen muy rápidamente. Se habrían conservado en eucariotas, porque presentan ventajas evolutivas; un mismo gen puede madurar de distintas maneras: 1 gen: distintas proteínas.
En los eucariotas el DNA está asociado a histonas, formando nucleosomas. Se ha observado que en genes que se transcriben continuamente (como los de RNAr) en los que el DNA está siempre extendido.
1.-Iniciación. Para la síntesis de ARNm existen dos señales de inicio, en una región del DNA llamada región promotora: la TATA, a 25 pares de bases del inicio de la transcripción hacia el extremo 5' y la CAAT algo más alejada. A partir de aquí empezará a transcribir. Esta región promotora indica dónde debe empezar y qué hebra va a leer.
-La RNA poli II reconoce y se une a la región promotora. Esta es una enzima que contiene una unidad sigma, que una vez comenzada la transcripción se suelta y es el resto el que va recorriendo el gen desde el extremo 5' utilizando como molde la otra hebra. Además hace que se abra la doble hélice.
-Desenrolla la doble hélice, una de cuyas cadenas servirá de molde
-La enzima se desplaza 3'---5' de la cadena de DNA molde y por tanto une nucleótidos que crecerán en dirección 5'---3'.Selecciona nucleótidos complementarios, que son triP , pasan a mP y eso les da E para catalizar la unión,
El proceso de síntesis continúa en sentido 5'-3'. Al cabo de 30 nucleótidos se añade una caperuza formada por una metil-guanosintriP al extremo 5'
-La ARNpoli une ribonucleótidos triP al OH 3' de la cadena de ARN con liberación de 2P. Puede comenzar la unión sin cebador
-Prosigue hasta la señal de terminación :TTATTT
-Para cada segmento de DNA sólo se transcribe una de las dos cadenas. ¿Quién determina cuál de la dos cadenas? La orientación del promotor
-Se traducen unos 30 nucleótidos por segundo, en eucariotas
-En eucariotas existen unas ARNpoli específicas para cada tipos de ARN :I, II y III
-Una vez terminada la cadena, sufrirá maduración
El promotor : TATA que lo reconoce la RNApoli II de tal forma que comienza 25 nucleótidos abajo y la CAAT algo más abajo
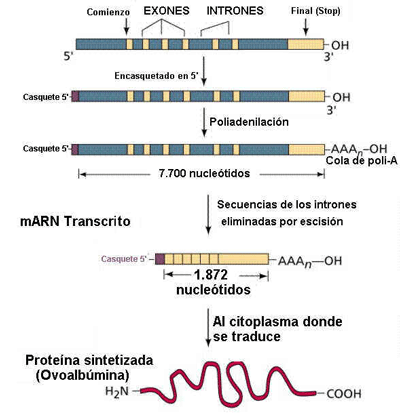
Elongación: cuando lleva 30 bases se le añade el RNAm en el extremo 5' una caperuza formada por metil guanosina triP, que sirve como lugar de anclaje al ribosoma y que aparece en eucariotas.
Terminación: su señal parece que es la secuencia TTATTT. La poliA polimerasa adiciona 150-200 nucleótidos de A que interviene en maduración y transporte del ARN fuera. En procariotas hay una secuencia palindrómica rica en G y C...TTTTT que hace que se forme bucle.
La mayor parte(hasta el 90%) del RNA es degradado
Maduración- En eucariotas, se realiza por medio de una enzima que corta los intrones (parece que los reconoce por GU.......AG) La lleva a cabo una enzima ribonucleopreína pequeña nuclear (RNPpn) que actúa al nivel del núcleo. Reconoce a los intrones, que suelen empezar por GU y acabar con AG los corta y los retira.
A continuación actúan RNA- ligasas específicas que empalman los exones.
Los RNAm que salen tienen 7.000 nucleótidos, mientras que en el citoplasma sólo tienen 1.500.Esto es debido al proceso de maduración que siguen los exones e intrones.
En procariotas se hace directamente.
2.-Traducción.
El RNAt se activa con ATP gracias a la aminoacil-RNAt-sintetasa. Estas enzimas acoplan cada Aa a su molécula de RNAt. Es el RNAt a través de su anticodón el que reconoce el codón del RNAm específico del Aa. Para cada Aa existen una sintetasa específica y esto supone el gasto de 1 ATP.
Hacen falta dos señales:
-Triplete iniciador AUG (que codifica la Met)
-Caperuza de metil guanosina en el RNAm
Comienza la lectura por el triplete más próximo a la caperuza
2.1.Iniciación: Unión del ribosoma (subunidad menor: con anticodón UAC) gracias a :
-Factores proteícos de iniciación }
-Energía del GTP } Complejo de iniciación + subunidad menor del ribosoma.
y lleva RNAt iniciador ( con Met) y anticodón complementario a AUG (todas tienen Met inicial que se puede eliminar). Al final se liberan FI y dejan paso a subunidad mayor del ribosoma que se acopla con complejo de iniciación formando el complejo activo con dos sitios de fijación el P y A libre para recibir a segundo RNAt con su Aa
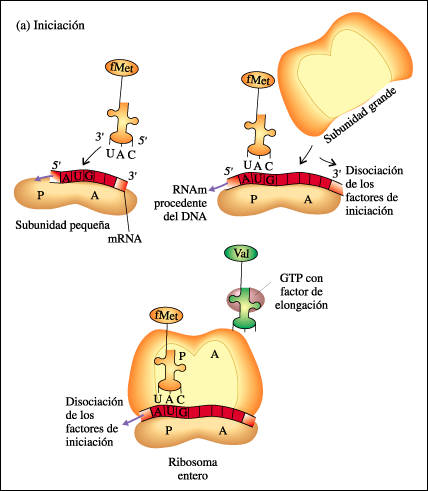
2.2.-Elongación de la cadena. Se inicia cuando un 2º Aminoacil ARNt con anticodón complementario entra en el ribosoma.El radicalcarboxilo del Aa iniciador se une con el amino del Aa siguiente mediante enlace peptídico.El RNAt sin Aa sale del ribosoma y se produce la translocación ribosomal. El ribosoma se desplaza a lo largo del RNAm en el sentido 5'-3' quedando libre de nuevo el sitio A
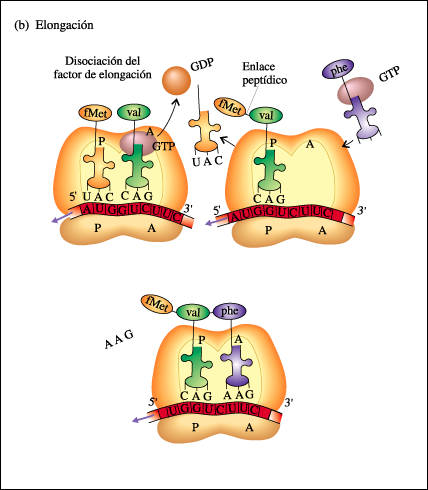
2.3.-Finalización de la síntesis.-Viene dada por los tripletes sin sentido:UAA, UAG y UGA.Son reconocidos por los factores proteicos de liberación, que se instalan sobre el A y provocan que se una el grupo COOH con agua, liberándose las cadena.
Un mismo RNAm puede ser traducido por varios ribosomas a la vez.
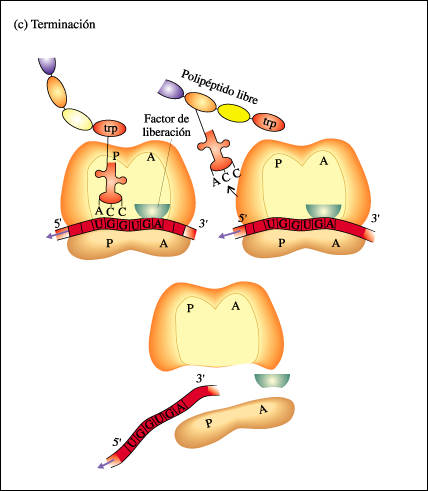
En procariotas la traducción es simultánea a la transcripción, formándose el típico arbollillo.
REGULACIÓN EN EUCARIOTAS
Se produce mediante activación de la transcripción de unos genes y la represión de otros.
El DNA de eucariotas se encuentra empaquetado densamente, por lo que suele resultar inaccesible al RNApoli.Para que una región que contiene determinados genes pueda transcribirse, tiene que ser previamente descondensada gracias a lo cuál, los nucleosomas se abren y adoptan una configuración extendida, desenrollándose las dos hebras.
¿Cómo se realiza la activación de la transcripción?
1.-Descondensación de la cromatina:frente a señales reguladoras del medio interno, generalmente hormonal, la cromatina se descondensa durante un período de tiempo corto, pero suficiente para que se transcriban los genes.La descondensación de cromatina se produce por modificaciones covalentes (acetilación y fosforilación) de determinados tipos de histonas, como la H1.
2.-Intervienen un conjunto de proteínas reguladoras que activan la transcripción de los genes localizados en la región de cromatina activa descondensada.Se cree que su activación responde a la acción de hormonas específicas.
Al parecer reconocen secuencias determinadas del DNA que presentan una conformación particular de la doble hélice: así, las regiones con doble hélice de configuración Z, podrían se zonas con genes inactivados, pues la hélice Z afectaría al promotor impidiendo que las proteínas activadoras favorezcan la fijación de RNApoli
En plantas, la luz actúa de activadora de genes vegetales, ya que controla el desarrollo de la planta mediante la fotomorfogénesis.Y además activa los fitocromos y estimula una secuencia reguladora del DNA próximo al promotor, actuándo como potenciadora
3.-Regulación hormonal. Gracias a los mecanismos homeostáticos, las células eucariotas no responden individualmente a las variaciones del ambiente externo, aunque si lo hacen frente al ambiente interno, como las hormonas.
×Hormonas esteróidicas(anabolizantes) y las tiroideas, atraviesan fácilmente la membrana plasmática por difusión y en el citoplasma se unen con proteínas receptoras.Este complejo se activa y a través de los poros del núcleo, se une a la cromatina y pone en marcha la transcripción de determinados genes.En el caso de anabolizantes se estimula la síntesis de proteínas musculares(culturista británica que se suicida, Ben Johnson descalificado...)Son derivados de la testosterona que estimula el crecimiento de la musculatura y potencia la actividad física. Suponen un esfuerzo para el hígado, dado que estimulan la síntesis de proteínas y provocan retención de líquidos.En EEUU se estima que unos 200.000 adolescentes lo están tomando, aunque ha descendido.Efectos:
En hombres: caída de cabello, aumento del tamaño de mamas, atrofia testicular.
En mujeres: aumento del vello corporal y voz más grave.
En ambos: trastornos mentales y aumento de agresividad, alta presión sanguínea y aumento de enfermedades coronarias, aumento de masa muscular y alteraciones en tendones, aumento de colesterol, detención del crecimiento óseo, anormalidades químicas en hígado....
×Hormonas proteicas, insulina, hormona del crecimiento, debido a su tamaño e hidrofilia no pueden atravesar la membrana, por lo que se une con un receptor específico de la membrana, se cataliza la transformación del ATP en AMPc, que actúa de segundo mensajero: activa las proteínas reguladoras de la transcripción y favorece la descondensación de la cromatina, por lo que se estimula la síntesis proteica.
El DNA como portador de información
El DNA de una sola célula humana que, si se extendiera en una hebra única mediría casi 2 metros de largo, puede contener una información equivalente a unas 600.000 páginas impresas de 500 palabras cada una, o a una biblioteca de aproximadamente 1.000 libros.
Una vez conocida la estructura del ADN se descifró el código en que está escrita la información genética.
La identificación del ARNm como copia de las instrucciones genéticas, no resolvía el cómo estaba codificada la información.
Las proteinas contienen 20 Aa distintos, pero los ácidos nucléicos sólo contienen 4 nucleótidos diferentes, por lo que:
-Si cada nucleótido codifica un Aa, sólo se especificarían 4 Aa.
- Si dos nucleótidos codifican un Aa, habría un máximo de 4x4 = sólo se especificarían 16 Aa.
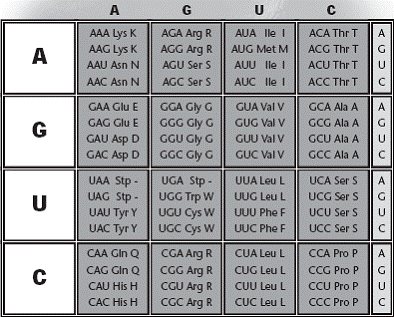
-Cómo mínimo hacen falta tres nucleótidos en la secuencia: 4x4x4 = 64 combinaciones posibles o codones, lo que basta para secuenciar 20 Aa.
En la década de 1960, se trató de deducir el código genético, es decir, el conjunto de instrucciones que determinan cómo será el organismo de cada especie. En esto se destacó el bioquímico norteamericano Marshall Nirenberg, quien en 1962 junto con Johann Heinrich Matthaei logró leer la primera palabra del alfabeto: encontró que cuando la molécula de ARN es UUU (contiene una secuencia de tres uracilos juntos), el organismo fabrica el aminoácido fenilalanina. Estos científicos lograron descifrar el código luego de realizar experimentos donde traducían distintos ARN mensajero (ARNm) compuestos por un único triplete repetido y analizaban el aminoácido producido.
Nirenberg y Matthaei colocaron 4 aminoácidos marcados radioactivamente en cada uno de los 5 tubos de ensayo. Luego agregaron enzimas, ribosomas, ATP y poliuracilo y notaron que en uno de los tubos de ensayo se había sintetizado grandes cantidades de un polipéptido desconocido. Para saber cuál aminoácido había producido ese polipétido, colocaron un solo aminoácido por tubo de ensayo: en el único tubo donde se sintetizó una proteína fue el que contenía fenilalanina.
Además, Nirenberg y sus colegas demostraron que el código es universal: los componentes del ADN son los mismos para reptiles, anfibios, bacterias y seres humanos.
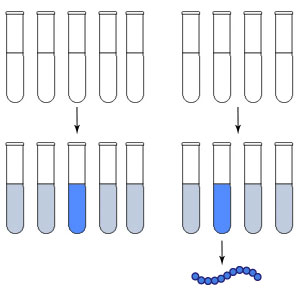
Así se fueron descubriendo los codones de ARNm para todos los aminoácidos.
Características de código genético:
• Tres nucleótidos sucesivos (del ARNm), constituyen un codón que codifica para
un aminoácido específico.
• Algunos aminoácidos pueden estar definidos por más de un codón. (Ver código
genético).
• 61 de las 64 posibles combinaciones de las tres bases nitrogenadas codifican
aminoácidos específicos.
• Existen 3 combinaciones que no codifican para aminoácidos UAA, UAG, UGA sino
que constituyen señales de terminación que indican que la cadena de proteína ha
terminado.
• El codón AUG codifica para el aminoácido metionina y adicionalmente funciona
como secuencia de iniciación.
Expondremos algunas de las características del código que son importantes para comprender las implicaciones patológicas de una mutación.
EL CÓDIGO GENÉTICO ESTÁ FORMADO POR TRIPLETES
-La estructura del ADN, la doble hélice, es independiente de la secuencia concreta de nucleótidos que la compone
EL CÓDIGO GENÉTICO ESTÁ FORMADO POR TRIPLETES
-La estructura del ADN, la doble hélice, es independiente de la secuencia concreta de nucleótidos que la compone
-La secuencia de nucleótidos es importante, no para la estructura, sino porque codifica una secuencia de aa que constituyen una proteína
-El código genético se lee en grupos de 3 nucleótidos denominados codones
-Un gen consta de una serie de codones que se leen secuencialmente desde un punto de inicio a uno de terminación.
Características del código genético
-NO ES AMBIGUO: Un triplete o codón sólo codifica un aminoácido
-ESTÁ DEGENERADO: Existen varios codones para el mismo aminoácido
-ES CASI UNIVERSAL: UGA en el ADN mitocondrial significa triptófano
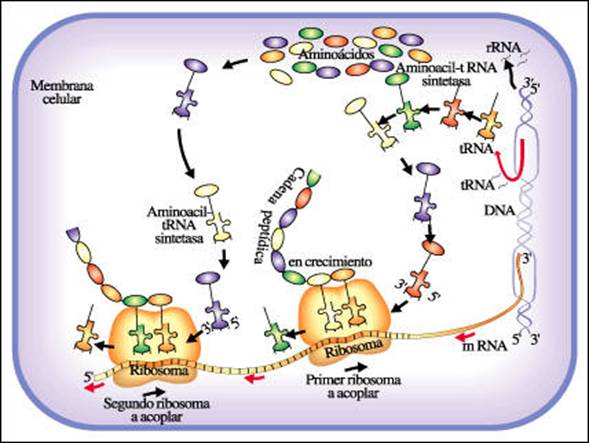
TRADUCCIÓN:
La síntesis de proteínas, más comúnmente conocida como traducción, es un proceso en el cual la información contenida en los genes (genotipo) es expresada en proteínas
(fenotipo). Tiene lugar en los ribosomas de manera similar en eucariotas y procariotas. Las reglas específicas para la traducción dependen del código genético, el lenguaje en el que se conserva la información que viene de los genes y es de
carácter universal.
El proceso de traducción tiene las siguientes etapas:
1. Iniciación: proceso de unión del ARNm a la unidad más pequeña del ribosoma, la
cual se encuentra libre en el citoplasma y se une en un extremo del ARNm exponiendo el codón iniciador, AUG (Adenina, Uracilo y Guanina) a la secuencia complementaria
en el ARNt (anticodón UAC). De este modo, las proteínas sintetizadas tendrán siempre al inicio el aminoácido metionina. A continuación se une la subunidad grande del ribosoma formándose así el ribosoma completo y funcional que tiene dos sitios claves:
- Sitio P (peptidil), ocupado por el ARNt-metionina
- Sitio A (aminoacil), libre para recibir un segundo ARNt cargado con un nuevo aminoácido.
2. Enlongación: consiste en la unión de los aminoácidos para la elaboración de la proteína. Cada vez que ARNt trae un aminoácido ocurre un proceso cíclico de
enlongación.
3. Terminación: fin de la síntesis de proteínas. La información de terminación está dada por los siguiente codones: UAA, UAG, UGA. Estos codones bloquean la unión del ARNt al ribosoma y hacen que se suelten las dos unidades del ribosoma y se libere la proteína.
Repasa estos conceptos
Transcripción |
Eliminación intrones |
Unión ARNm al ribosoma |
ARNt se une al Aminoácido |
Traducción: uniones sucesivas de ARNt portando Aa |
Interrupción de síntesis polipeptídica |
Formación de la proteína. |
Regulación de la expresión de los genes
Para que toda secuencia que codifica en el ADN (exones) se exprese en proteínas, es necesario que exista un proceso de control conocido como regulación genética.
En procariotas la regulación genética ocurre a través de operones. Un operón es una asociación de genes cada una de las cuales codifica una proteína.
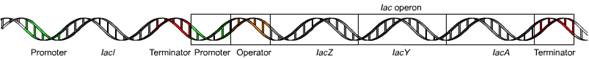
El operón consta de un promotor, una región de control y una secuencia de genes codificantes.
La expresión o no de las secuencias codificantes dependerá esencialmente de la
disponibilidad de las proteínas codificadas en el entorno. La expresión de las proteínas está regulada por la unión de una proteína (promotor o represor) al promotor o control.
- Genes estructurales: catalizan la reacción de hidrólisis de la lactosa en glucosa más galactosa, facilitan el transporte de β-galactósidos al interior de la bacteria y catalizan la transferencia del grupo acetil del acetil Coenzima A respectivamente
- Gen lac z:
- Gen lac y:
- Gen lac a:
- Promotor: reconoce la RNA polimerasa para llevar a cabo la transcripción.
- Operador: es reconocida por la proteína represora Lac I.
- Gen represor (lac I): Impide la transcripción de los genes bajo el control de este promotor pero estimula la unión de la RNA polimerasa formando lo que se conoce como Complejos Cerrados. Cuando el represor se retire (en presencia de inductor que en este caso será lactosa o IPTG), la RNA polimerasa estará lista para formar Complejos Abiertos y empezar la transcripción.
En Eucariotas se han definido los siguientes elementos básicos de regulación:
1. Los promotores: Son regiones en el ADN que señalan el inicio de la secuencia
codificadora y a las que se unen los factores de transcripción y la polimerasa. El
promotor más importante es la caja TATA, una corta secuencia de pares de bases T-A
y A-T.
2. Enhacers: regiones fomentadores de la transcripción.
3. Silenciadores: bloquean la síntesis de proteínas cuando existe sobreexpresión, es
decir cuando los niveles de síntesis para una proteína exceden el nivel normal.
La regulación genética consiste en encender y apagar los genes.
Actualizado en abril de 2009
![]()