
Anabolismo
-Concepto y tipos
-Anabolismo autótrofo
-Fotosíntesis. Estudio detallado.
-Concepto y función biológica.
-Estructuras que intervienen en la fotosíntesis. Ubicación :Cloroplastos, Pigmentos fotosintéticos, Fotosistemas.
-Fases de la fotosíntesis
-Sustratos necesarios, productos finales y balance energético total.
-Importancia biológica y medioambiental.
-Comparación con la fotosíntesis bacteriana.
Quimiosíntesis. Concepto e importancia biológica del proceso.
-Anabolismo heterótrofo
Concepto y tipos
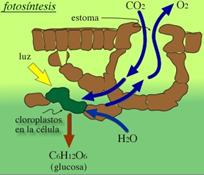
Mientras una célula heterótrofa cataboliza moléculas de glucosa procedentes del medio, una autótrofa debe sintetizar previamente la glucosa antes de poderla catabolizar. El proceso de síntesis de biomoléculas que parte del dióxido de carbono y del agua como sustratos iniciales y requiere como fuente de energía la luz solar, recibe el nombre de fotosíntesis.
Gracias a la fotosíntesis nos alimentamos, respiramos, nos vestimos y hacemos funcionar los motores.
La energía solar, a pesar de ser la más abundante de la Tierra, no está siempre disponible para las plantas, sino solamente de día.¿Qué hacen las plantas para disponer de energía también durante la noche?
Cuando las células disponen de luz-reacciones luminosas- la utilizan para producir moléculas de ATP y NADPH.En ausencia de luz-reacciones oscuras- las células utilizan ATP y NADPH para sintetizar glucosa, que almacenan en forma de almidón.
En cualquier momento del día o de la noche, la planta puede degradar el almidón, mediante la respiración, y obtener energía.
Puedes comenzar con este repaso
Si quieres saber más sobre el ciclo del CO2
Breve historia de la fotosíntesis
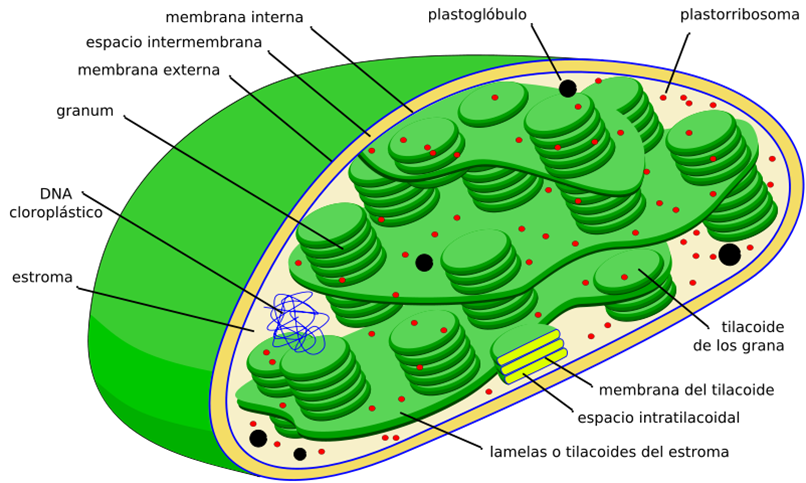
Todas las células fotosintetizadoras, a excepción de bacterias y cianobacterias tienen cloroplastos. En estos orgánulos están presentes las moléculas de clorofila. Cada molécula de estos pigmentos de color verde contiene un anillo porfirínico con un átomo de Mg.
En gral, cuando una molécula absorbe luz, sus electrones son impulsados a un nivel energético superior. Normalmente, esta energía adicional es disipada en forma de luz o calor y los electrones retornan a su estado inicial. Sin embargo, en moléculas como la clorofila, cuando absorben la energía de la luz, sus electrones (los que mantienen el enlace del átomo de Mg con el anillo porfirínico) son excitados y pueden cederse fácilmente a un aceptor.
Los electrones excitados son recogidos por proteínas transportadoras, algunas de las cuales asocian el transporte electrónico a un bombeo de Hidorgeniones del estroma del cloroplasto al interior del espacio tilacoide; la vuelta de los H al estroma se acopla a la síntesis de aTP de forma similar a lo que ocurre en la mitocondria. Este proceso de formación de ATP, impulsado por la luz se denomina fotofosforilación o fotofosforilación fotosintética.
Al final del transporte electrónico, puede producirse la reducción del nucleótido NADP.
El ATP y NADPH formados pueden utilizarse en las reacciones bioquímicas necesarias para reducir el dióxido de carbono y obtener biomoléculas orgánicas, pero puede usarse también para otras actividades vitales.
Por esto se ha considerado la fotosíntesis como un proceso de dos fases, cada una de las cuales tiene sus reacciones químicas características. Es proceso de excitación electrónica de la clorofila y obtención del aTP y NADP es la fase fotoquímica o lumínica.
La 2ª etapa es la fase oscura, con la reducción del dióxido y su incorporación a las cadenas carbonadas. Las reacciones, se denominan ciclo de Calvin
Concepto y función biológica.
- Las células autótrofas fabrican su alimento a partir de sustancias inorgánicas sencillas. La mayoría utilizan luz solar y al proceso se le denomina fotosíntesis.
- Se realiza en los cloroplastos
- Lo hace en dos fases:
- Luminosa
- Oscura
- Es el proceso a través del cuál los organismos productores, a partir de CO2, del agua, sales minerales y luz solar fabrican su propia materia
- Se transforma así la energía lumínica en energía química.
- Para ello: Todas las célula fotosintetizadoras, menos las bacterias, tienen cloroplastos
Cómo se ven los cloroplastos en Elodea
Fase luminosa Con ella, se obtiene ATP y NADPH, gracias a la excitación de la clorofila |
Fase oscura |
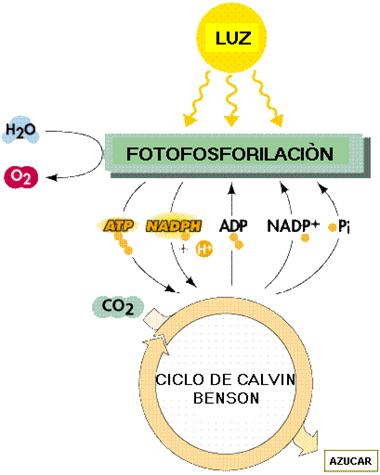
Esquema fotosíntesis
Extraído de Anaya
Fase luminosa
1.-Naturaleza de la luz
La luz son ondas electromagnéticas emitidas por el sol y con movimiento ondulatorio.En sus interacciones con la materia se comporta como si estuviera compuesta de pequeños paquetes de energía- quantos-denominados fotones.
El espectro electromagnético de la radiación emitida por el Sol ha sido dividido arbitrariamente en regiones de diferente longitud de onda:
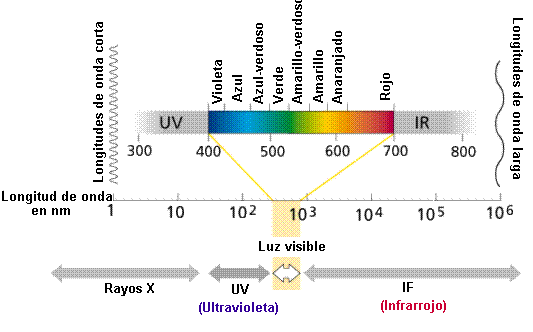
Modificado de: http://www.whfreeman.com/life/update/.
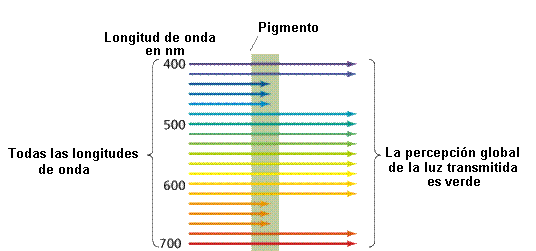
La luz visible es una pequeña parte del espectro electromagnético. Cuanto más larga la longitud de onda de la luz visible tanto más rojo el color. Asimismo las longitudes de onda corta están en la zona violeta del espectro. Las longitudes de onda mas largas que las del rojo se denominan infrarrojas, yaquellas mas cortas que el violeta, ultravioletas.
Las radiaciones altamente energéticas (Rayos gamma, rayos X y rayos uv) producen ionización de la materia, sin embargo, la luz visible está formada por un conjunto de radiaciones entre 400 y 700 nm que son de bajo contenido energético.
- En ella, tiene lugar la reacción:
CO2 + NADP+ ½ O2 + NADPH + H+
Como este proceso no es espontáneo, hace falta energía, aportada por la luz y transformada en energía química por la clorofila.
2.-Los pigmentos fotosintéticos
- Son las sustancias capaces de captar la energía solar en la fotosíntesis.Son moléculas que absorben luz de una determinada longitud de onda y por eso son coloreadas. El color del pigmento esta dado por la longitud de onda no absorbida (y por lo tanto reflejada). Los pigmentos negros absorben todas las longitudes de onda que les llega y los pigmentos blancos reflejan prácticamente toda la energía que les llega. Los pigmentos tienen un espectro de absorción característico de cada uno de ellos.
- Las clorofilas constan de cuatro anillos de pirrol unidos por medio de puentes de metilo (--CH=) lo que constituye una porfirina. El tetrapirrol es el cuerpo básico de las porfirinas, dentro de las cuales se incluyen además de las clorofilas, las hemoglobinas y los citocromos. La característica cromófora de la clorofila se debe al sistema de dobles enlaces conjugados generados por la unión de los anillos de pirrol mediante los grupos metino. En el centro del sistema de anillos se halla un átomo metálico: para las clorofilas es el magnesio, para la hemoglobina el hierro.
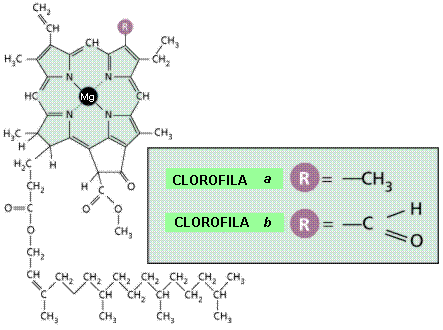
- Las clorofilas absorben todos los colores del espectro excepto el verde, y a eso se debe el color verde.La clorofila se suele degradar en otoño para recuperar el nitrógeno y magnesio que posee antes de que pierdan las hojas.
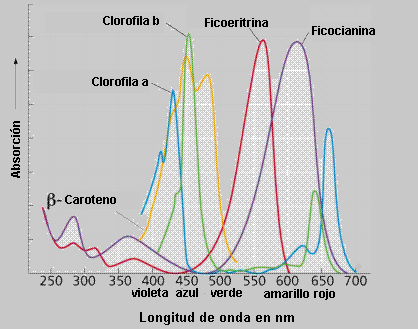
- Los pigmentos accesorios que incluyen a la clorofila b (también c, d, y e en algas y protistas) y los carotenoides, como el beta caroteno, son pigmentos rojos, anaranjados o amarillos, que en las hojas verdes están enmascarados por las clorofilas, que son más abundantes; sin embargo en algunos tejidos, como los del tomate maduro, predominan los colores reflejados por los carotenoides. Y las xantófilas (carotenoide de color amarillo), absorben la energía no absorbida por la clorofila, por lo que el conjunto de todos los pigmentos abarca casi todo el espectro.La energía captada debe transmitirse a moléculas de clorofila a , que son las que conectan con los sistemas de transporte electrónico.
- Los carotenoides y la clorofila b absorben en la longitud de onda del verde. Ambas clorofilas también absorben en la región final del espectro (anaranjado - rojo), o sea longitudes de onda larga y menor cantidad de energía.
El origen de los organismos fotosintéticos en el mar da cuenta de esto. Las ondas de luz mas cortas (y de mayor energía) no penetran mas allá de los 5 metros de profundidad en el mar. La habilidad para obtener energía de las ondas mas largas (y penetrantes en este caso) pudo constituir una ventaja para las primeras algas fotosintéticas que no podían permanecer en la zona superior del mar todo el tiempo. Explica también la disposición espacial de algas verdes, rojas y pardas: en superficie habría algas verdes en busca de la radiación roja, más abajo las algas pardas y por fin, las algas rojas en zonas profundas de dominio de la radiación verde, aunque hoy se ha comprobado que influye más la intensidad de la luz.
- Las plantas evolucionaron para utilizar los electrones del agua y transferirlos a transportadores de electrones, como el NADPH, para realizar reacciones bioquímicas benéficas. El problema es que un solo fotón no contiene suficiente energía para elevar un electrón al nivel de energía requerido en una sola etapa. Por tal razón, las plantas utilizan dos diferentes fotosistemas que funcionan en serie para elevar el nivel de energía de los electrones mediante la captura de dos fotones consecutivos .
- Cada fotosistema está formado por un conjunto de pigmentos antena y un centro de reacción, que tiene la capacidad de realizar las reacciones fotoquímicas necesarias. Los centros de reacción en estos fotosistemas son pigmentos específicos que han sido denominados P-680 y P-700. Por ello, los pigmentos se agrupan formando los fotosistemas (son el conjunto de la clorofila diana, el dador de electrones, el complejo antena y el aceptor de electrones), que se localizan en las membranas de los tilacoides.
- En cada fotosistema, los pigmentos se distribuyen de manera que actúan como antenas o colectores, absorbiendo la energía luminosa y transmitiéndola al llamado centro de reacción, que contiene una molécula especial de clorofila a. Se asocian a determinadas proteínas comportándose como una antena solar, cada una de ellas es una trampa fotónica, donde varios centenares (unos 300) de pigmentos fotosensibles (que captan longitudes de onda menor) y proteínas se disponen de forma que la energía lumínica absorbida por cada una de estas moléculas se canaliza por medio de proteínas hacia una molécula de clorofila (Que capta energía de mayor longitud de onda).
- Así, cada antena, se comporta como un embudo destinado a recoger la energía fotónica de la luz y encauzarla hacia los componentes fotoactivos del centro de reacción, que son dímeros de clorofila a (P-700 y P-680) y forman parte de los fotosistemas I y II.
En las membranas tilacoidales de los cloroplastos se encuentran los fotosistemas
Resumen
| H2O + NADP+ | NADPH + H+ + O2 |
Por cada electrón que fluye se absorben dos fotones, uno en cada fotosistema.
Fase luminosa
Observa la animación sobre la actuación de fotosistemas:
http://croptechnology.unl.edu/animationOut.cgi?anim_name=fotosintesis_spanish.swf, de la Universidad de Nebraska
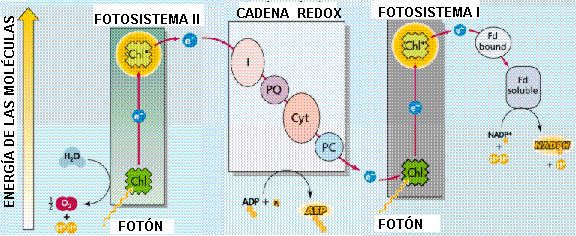
1.-Son reacciones que captan energía: transforman la energía solar en química. Como resultado, se sintetiza ATP y la reducción de los coenzimas celulares, NADP a NADPH. Tiene lugar en la membrana de los tilacoides de los cloroplastos e intervienen en ella los fotosistemas. Se habla de Fotofosforilación no cíclica y la cíclica
Teoría quimiósmotica
Modificada de: http://www.whfreeman.com/life/update/.
Fase oscura

2.-Reacciones de fijación del carbono. Formado por el ciclo de Calvin. En ellas, el ATP y el NADPH producidos en la primera fase, se utiliza como fuente de energía y poder reductor para transformar sustancias inorgánicas oxidadas (CO2, NO3, SO4) en moléculas orgánicas reducidas que participarán en la síntesis de moléculas orgánicas complejas.
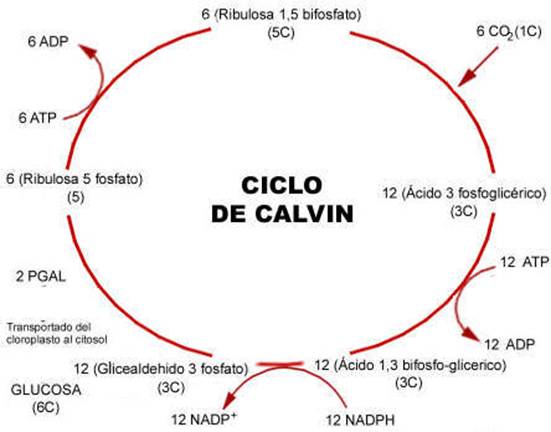
Las reacciones se producen en el estroma del clorplasto y en el proceso intervienen diversas moléculas.
Comienza al reaccionar la ribulosa 1-5-difosfato con el CO2. Este sale como gliceraldehido-3-P. En cada vuelta del ciclo se consumen 3 ATP y 2 NADPH, siendo necesarias 3 vueltas del ciclo para formar una molécula de gliceraldehido-3-P
Aunque la mayoría de las plantas siguen este modelo, en zonas de clima tropical, como adaptación a altas temperaturas y escasez de agua, plantas como la caña de azúcar y la remolacha, desarrollan otras reacciones. El proceso es también cíclio, per la primera molécula resultante es un compuesto de 4 C, por lo que a estas plantas se les denomina plantas C4, en oposición a las que realizan Calvin, que son C3
Balance energético de la fotosíntesis: La fase luminosa de la fotosíntesis produce ATP y NADPH poder reductor, aprovechable por otras reacciones) . Si se sintetiza una molécula de glucosa (C6H12O6) se necesitan 6 CO2 y 12 de Agua. El agua libera 6 O2 a la atmósfera y aporta 12 hidrógenos de la glucosa y los 12 hidrógenos necesarios para pasar los 6 O2 sobrantes del CO2 a Agua. Intervienen 24 Hidrógenos. Aparecen así 24 protones y 24 electrones y, como cada electrón precisa dos fotones (uno en el PSI y otro en el PSII), se necesitan 48 fotones. El ciclo de Calvin necesita por cada CO2 incorporado, 2 NADPH y 3 ATP. Para una molécula de glucosa se necesitan 12 NADPH y 18 ATP.
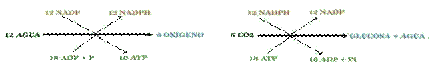
Balance energético del ciclo de Calvin
Como resultado de las reacciones del ciclo de Calvin se produce la reducción del
dióxido de carbono, que se incorpora a la vía del anabolismo de los glúcidos. El primer
metabolito que conduce a la síntesis de glucosa es el gliceraldehído-3-fosfato
(G3P) que se obtiene en el ciclo de Calvin. Son necesarias tres vueltas del ciclo para
sintetizar una molécula de G3P. El proceso puede resumirse mediante la siguiente
ecuación:
| 3 Ribu-1,5-di-P + 3CO2 + 6(NADPH + H) + 9 ATP | 3Ribu-1,5,di-P + G3P + 6 NADP+ + 9ADP + Pi |
FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS
La actividad fotosintética varía en función de diversos factores que influyen en su
rendimiento y que puedes ver a continuación.
Variación de la actividad fotosintética en función de ciertos factores
1.- Intensidad de la luz
La fotosíntesis es proporcional a la intensidad de la luz: a mayor intensidad, mayor
es la tasa de fotosíntesis, hasta un punto en que la actividad se mantiene constante.
Concentración de CO2
El CO2 es necesario en las reacciones del ciclo de Calvin y, en condiciones normales,
es el factor limitante más importante de la fotosíntesis. Un aumento de su concentración
produce un aumento de la fotosíntesis hasta un valor de asimilación máximo
que depende de cada organismo.
En la gráfica se muestra la influencia conjunta de la intensidad luminosa y la concentración de CO2.
Concentración de O2
Al aumentar la concentración de O2 disminuye el rendimiento de la fotosíntesis; esto
es debido a la competencia que se establece entre el O2 y el CO2, favoreciéndose el
proceso de la fotorrespiración.
Temperatura
En la fase lumínica no influye de forma importante la temperatura. La actividad de
los enzimas sigue la curva típica de mayor actividad a mayor temperatura.
Agua y humedad
El agua no es un factor limitante de la fotosíntesis, salvo en condiciones excepcionales.
La humedad influye en la apertura y cierre de estomas. Si el ambiente es seco,
los estomas se cierran y se impide así el intercambio de gases imprescindible para la
fotosíntesis. Esto puedes verlo en la siguiene animación.
Animación: Apertura y cierre de estomas
La apertura y el cierre de los estomas se produce por el siguiente mecanismo:
Durante el día, las células oclusivas de los estomas realizan la fotosíntesis. El aumento
de la presión osmótica motivado por este proceso produce la entrada de agua en
ellas y se hacen más turgentes. La zona de su pared que da al ostiolo, que es más
gruesa, se curva y el estoma se abre.
Por la noche no se produce fotosíntesis, las células oclusivas pierden turgencia y el
estoma se cierra.
Factores que regulan la producción primaria
Hay una serie de factores limitantes o reguladores que limitan la producción de los
autótrofos. De ellos pueden destacarse cinco: la luz y la organización del aparato fotosintético,
la cantidad de CO2, el agua, la disponibilidad de fósforo y nitrógeno y la
temperatura.
– La luz es imprescindible para el crecimiento vegetal. Según aumenta su intensidad,
aumenta la producción primaria; pero llega un momento en que el aparato fotosintético
se satura, y sucesivos aumentos de luz ya no se corresponden con aumentos
en la producción. Ello se debe a que existe una limitación intrínseca en el
propio orgánulo de captación de luz por dos causas:
• La disposición de las moléculas de clorofila en los cloroplastos, que se dan
“sombra” unas a otras.
• Que los centros de reacción que transforman la luz en energía química se encuentran
en número muy inferior a las moléculas de pigmento.
Así se justifica que un incremento en la intensidad de luz, por encima de un cierto
nivel, no lleve aparejado el aumento de la fotosíntesis. El mecanismo actúa como un
“cuello de botella” en el que, a partir de una cierta cantidad de agua, no podrá entrar
más al recipiente, aunque el flujo de esta siga aumentando. Esto explica que los
máximos rendimientos fotosintéticos se logren en las primeras horas de la mañana y
del atardecer, cuando la luz es más difusa, y no en las horas centrales del día, con
luz solar directa.
La selección natural ha actuado en el sentido de favorecer una cantidad mínima de
producción a intensidades luminosas bajas, sufiente para asegurar la permanencia
de los ecosistemas, y no en el de lograr la máxima conversión de energía posible.
Por ello son dudosas las esperanzas de elevar la producción de cultivos y así aumentar
los alimentos para la humanidad.
En relación con la energía radiante que llega a la Tierra, el rendimiento medio en
biomasa de la fotosíntesis es de aproximadamente el 1%. Sin embargo, algunas plantas,
como el maíz o la caña de azúcar, tienen una eficiencia fotosintética tres o cuatro
veces superior que la de otras plantas cultivadas.
– Una concentración de CO2 baja en el aire es un factor limitante para la producción
de materia orgánica. La asimilación de CO2 aumenta hasta que se estabiliza
cuando se alcanza una concentración del gas cuatro veces superior a la del aire
(entre 0,3 y 1,2 por mil). Por ello, algunas plantas cultivadas en invernaderos con
atmósfera enriquecida en CO2 aumentan netamente su productividad.
Sin embargo, otras plantas cultivadas, como la caña de azúcar, el maíz, el mijo y el
sorgo, crecen muy rápido con pequeñas concentraciones de CO2 en el aire y no aumentan
su crecimiento cuando se aumenta la cantidad de gas. Son las llamadas plantas
C4, frente a las restantes, a las que se denomina C3. Las siglas C3 y C4 se refieren
al número de átomos de carbono que presenta el primer compuesto que resulta de
la fijación sintética del CO2. El mecanismo más evolucionado es el C4. Las plantas C4
son, en su mayor parte, propias de regiones tropicales secas, con fuerte insolación,
temperaturas altas y humedad baja. Aunque fijan en el parénquima clorofílico de sus
Unidad 13. El anabolismo. La fotosíntesis y la quimiosíntesis
hojas, que se encuentran formando vainas que rodean a los vasos conductores, alejado
de la epidermis.
Otro tipo de plantas, las CAM (Crasulaceam Acid Metabolism), como son las cactáceas,
crasuláceas y algunas euforbias, están morfológica y fisiológicamente adaptadas a
condiciones desérticas. Realizan la fijación del CO2 por la noche, lo que les permie
tener cerrados los estomas durante el día, con el consiguiente ahorro de agua.
– La falta de agua en los vegetales terrestres impide que pueda llevarse a cabo el
proceso fotosintético. Por debajo de una cierta cantidad, los estomas se cierran,
impidiendo la entrada de CO2, con lo que disminuye el rendimiento fotosintético.
Por otra parte, en las plantas terrestres, al ser el agua el vehículo para las sales minerales,
si no hay un aporte suficiente, no habrá síntesis orgánica. Por ello se considera
que el crecimiento de un vegetal es un porcentaje determinado del agua que ha circulado
a través de la planta y se ha perdido a través de las hojas.
– El nitrógeno y el fósforo son tan esenciales para la síntesis de materia orgánica como
el CO2 y el agua. La proporción media entre los átomos de carbono, nitrógeno
y fósforo en la materia viva es de 106:16:1. Dado que en el entorno la proporción
está desviada a favor del carbono, frente al fósforo, la cantidad de fósforo es el
factor más importante que limita la producción primaria en la biosfera.
El nitrógeno tiene una importancia menor, debido a la gran cantidad que, en forma
de gas, se encuentra en la atmósfera y disuelto en el agua. Si falta nitrógeno en el
entorno, intervienen los organismos fijadores de nitrógeno gaseoso o moléculas
(bacterias y cianobacterias), que lo incorporan al ciclo del ecosistema en forma de
amonio, nitritos o nitratos.
Las actividades humanas movilizan grandes cantidades de fósforo, que es el componente
fundamental de los abonos agrícolas. De esta manera, una fracción importante
queda inmovilizada en forma de compuestos muy insolubles, perdiéndose para el
ciclo global del nutriente. Los ecosistemas naturales, como los bosques, se comportan
de forma diferente, ya que reciclan el fósforo con pérdidas mínimas, si es que las
hay.
– Una temperatura alta produce, si los otros factores están disponibles, un crecimiento
más rápido de los vegetales. Sin embargo, la respiración puede aumentar
más que la PPB, con lo que la producción neta no será necesariamente más alta a
mayor temperatura.
LA QUIMIOSÍNTESIS
Los organismos capaces de sintetizar materia orgánica a partir de compuestos inorgánicos
son autótrofos. Los fotosintéticos utilizan la luz como fuente de energía necesaria
para llevar a cabo el proceso. Otros, los quimiolitótrofos, realizan la quimiosíntesis.
La quimiosíntesis consiste en la obtención de materia orgánica a partir de
inorgánica y de la energía desprendida de reacciones químicas redox exergónicas.
Los principales organismos quimiliotótrofos y sus características se desarrollan a
continuación.
Las bacterias que realizan la quimiosíntesis
• BACTERIAS NITRIFICANTES
– Nitrosomas: Oxidan compuestos que contienen amoniaco en disolución que se
encuentra como ión amonio a nitrito.